

Stressor exposure disrupts commensal microbial populations in the intestines and leads to increased colonization by Citrobacter rodentium
- PMID: 20145094
- PMCID: PMC2849416
- DOI: 10.1128/IAI.00862-09
Stressor exposure disrupts commensal microbial populations in the intestines and leads to increased colonization by Citrobacter rodentium
Abstract
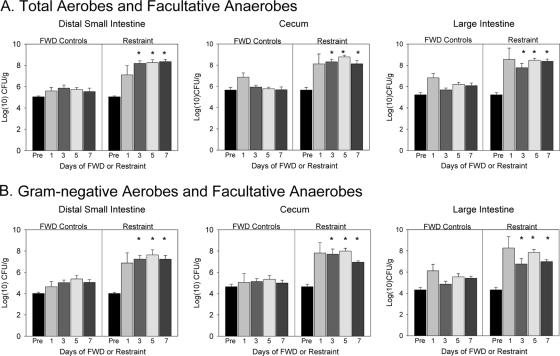
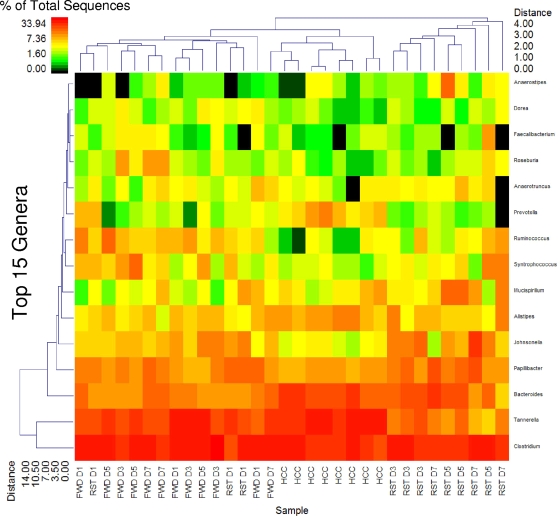
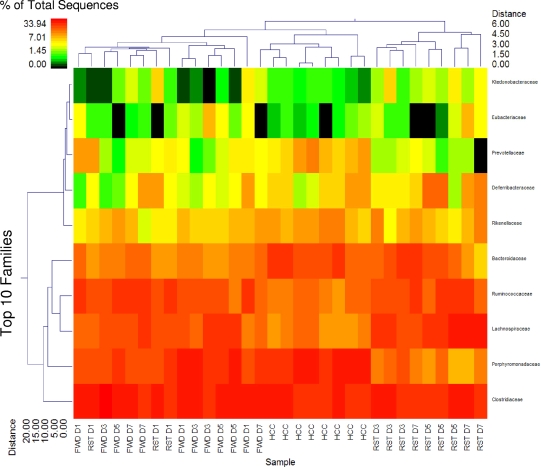
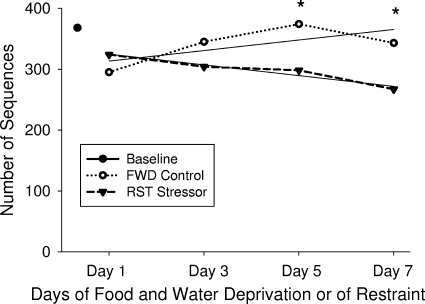
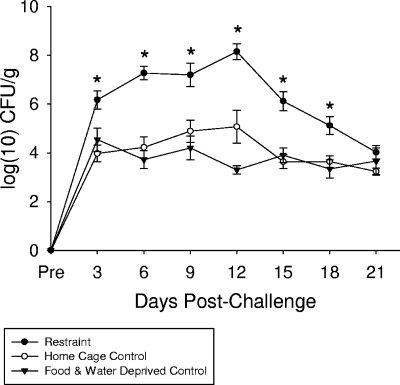
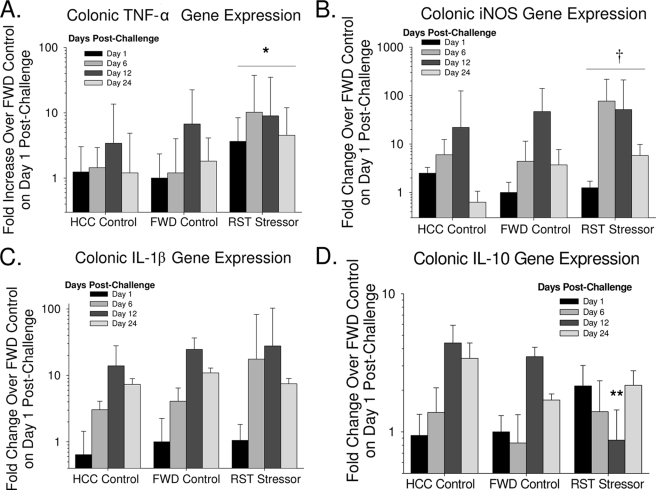
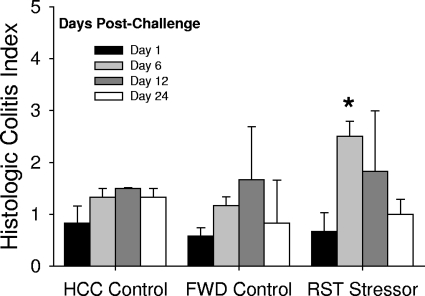
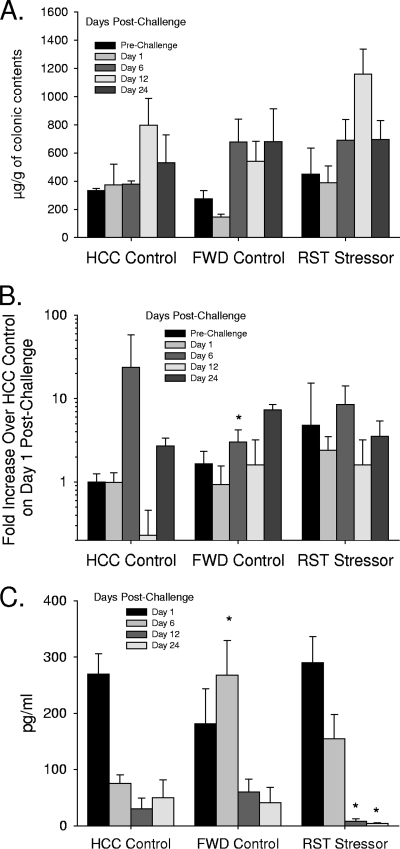
The gastrointestinal tract is colonized by an enormous array of microbes that are known to have many beneficial effects on the host. Previous studies have indicated that stressor exposure can disrupt the stability of the intestinal microbiota, but the extent of these changes, as well as the effects on enteric infection, has not been well characterized. In order to examine the ability of stressors to induce changes in the gut microbiota, we exposed mice to a prolonged restraint stressor and then characterized microbial populations in the intestines using both traditional culture techniques and bacterial tag-encoded FLX amplicon pyrosequencing (bTEFAP). Exposure to the stressor led to an overgrowth of facultatively anaerobic microbiota while at the same time significantly reducing microbial richness and diversity in the ceca of stressed mice. Some of these effects could be explained by a stressor-induced reduction in the relative abundance of bacteria in the family Porphyromonadaceae. To determine whether these alterations would lead to increased pathogen colonization, stressed mice, as well as nonstressed controls, were challenged orally with the enteric murine pathogen Citrobacter rodentium. Exposure to the restraint stressor led to a significant increase in C. rodentium colonization over that in nonstressed control mice. The increased colonization was associated with increased tumor necrosis factor alpha (TNF-alpha) gene expression in colonic tissue. Together, these data demonstrate that a prolonged stressor can significantly change the composition of the intestinal microbiota and suggest that this disruption of the microbiota increases susceptibility to an enteric pathogen.
Figures
Similar articles
-
The structures of the colonic mucosa-associated and luminal microbial communities are distinct and differentially affected by a prolonged murine stressor.Gut Microbes. 2014;5(6):748-60. doi: 10.4161/19490976.2014.972241. Gut Microbes. 2014. PMID: 25536463 Free PMC article.
-
The commensal microbiota exacerbate infectious colitis in stressor-exposed mice.Brain Behav Immun. 2017 Feb;60:44-50. doi: 10.1016/j.bbi.2016.09.010. Epub 2016 Sep 12. Brain Behav Immun. 2017. PMID: 27633986 Free PMC article.
-
Stressor exposure has prolonged effects on colonic microbial community structure in Citrobacter rodentium-challenged mice.Sci Rep. 2017 Mar 27;7:45012. doi: 10.1038/srep45012. Sci Rep. 2017. PMID: 28344333 Free PMC article.
-
The contributing role of the intestinal microbiota in stressor-induced increases in susceptibility to enteric infection and systemic immunomodulation.Horm Behav. 2012 Aug;62(3):286-94. doi: 10.1016/j.yhbeh.2012.02.006. Epub 2012 Feb 15. Horm Behav. 2012. PMID: 22366706 Review.
-
Overview of the Effect of Citrobacter rodentium Infection on Host Metabolism and the Microbiota.Methods Mol Biol. 2021;2291:399-418. doi: 10.1007/978-1-0716-1339-9_20. Methods Mol Biol. 2021. PMID: 33704766 Review.
Cited by
-
The structures of the colonic mucosa-associated and luminal microbial communities are distinct and differentially affected by a prolonged murine stressor.Gut Microbes. 2014;5(6):748-60. doi: 10.4161/19490976.2014.972241. Gut Microbes. 2014. PMID: 25536463 Free PMC article.
-
Intestinal microbiota, probiotics and mental health: from Metchnikoff to modern advances: part III - convergence toward clinical trials.Gut Pathog. 2013 Mar 16;5(1):4. doi: 10.1186/1757-4749-5-4. Gut Pathog. 2013. PMID: 23497650 Free PMC article.
-
Gut dysbiosis impairs recovery after spinal cord injury.J Exp Med. 2016 Nov 14;213(12):2603-2620. doi: 10.1084/jem.20151345. Epub 2016 Oct 17. J Exp Med. 2016. PMID: 27810921 Free PMC article.
-
Stress-Induced Neurodegeneration: The Potential for Coping as Neuroprotective Therapy.Am J Alzheimers Dis Other Demen. 2020 Jan-Dec;35:1533317520960873. doi: 10.1177/1533317520960873. Am J Alzheimers Dis Other Demen. 2020. PMID: 32969239 Free PMC article.
-
Mice Deficient in Epithelial or Myeloid Cell Iκκβ Have Distinct Colonic Microbiomes and Increased Resistance to Citrobacter rodentium Infection.Front Immunol. 2019 Sep 10;10:2062. doi: 10.3389/fimmu.2019.02062. eCollection 2019. Front Immunol. 2019. PMID: 31552024 Free PMC article.
References
-
- Acosta-Martinez, V., S. Dowd, Y. Sun, and V. Allen. 2008. Tag-encoded pyrosequencing analysis of bacterial diversity in a single soil type as affected by management and land use. Soil Biol. Biochem. 40:2762-2770.
-
- Bailey, M., H. Engler, J. Hunzeker, and J. F. Sheridan. 2003. The hypothalamic-pituitary-adrenal axis and viral infection. Viral Immunol. 16:141-157. - PubMed
-
- Bailey, M. T., and C. L. Coe. 1999. Maternal separation disrupts the integrity of the intestinal microflora in infant rhesus monkeys. Dev. Psychobiol. 35:146-155. - PubMed
-
- Bailey, M. T., G. R. Lubach, and C. L. Coe. 2004. Prenatal stress alters bacterial colonization of the gut in infant monkeys. J. Pediatr. Gastroenterol. Nutr. 38:414-421. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
