

Role of brain glycogen in the response to hypoxia and in susceptibility to epilepsy
- PMID: 26578889
- PMCID: PMC4621300
- DOI: 10.3389/fncel.2015.00431
Role of brain glycogen in the response to hypoxia and in susceptibility to epilepsy
Abstract
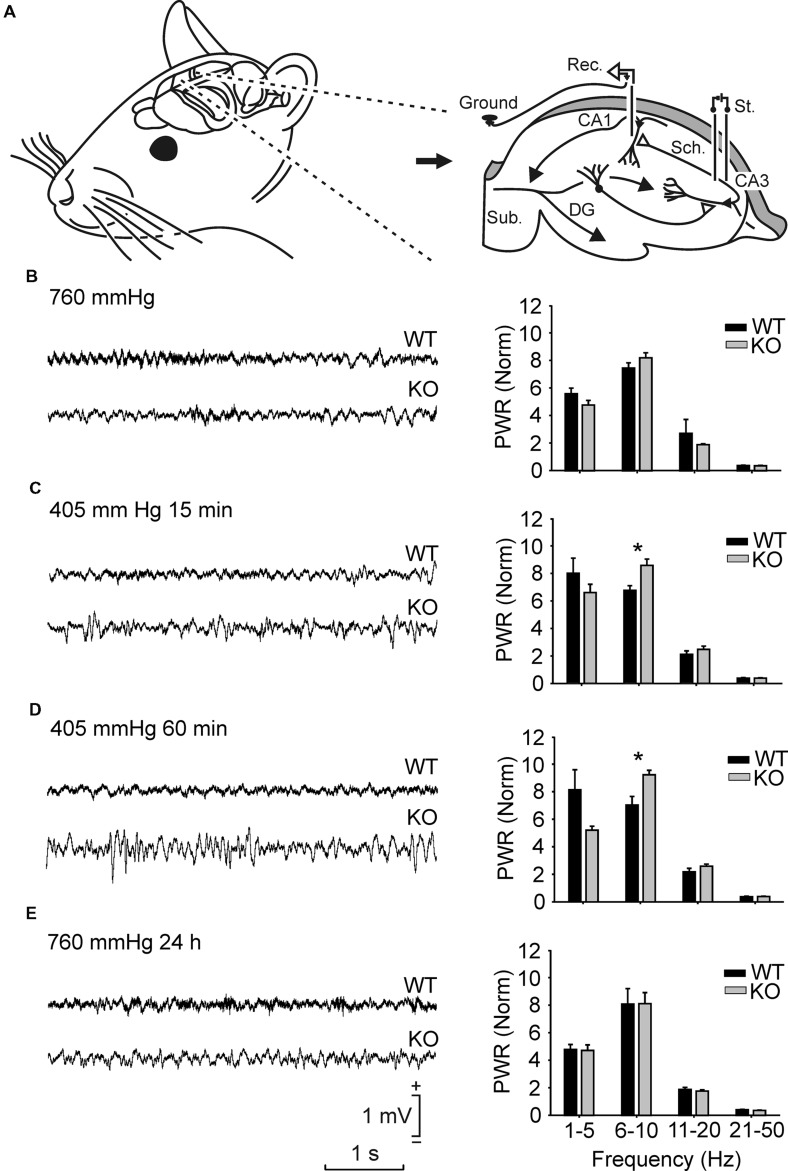
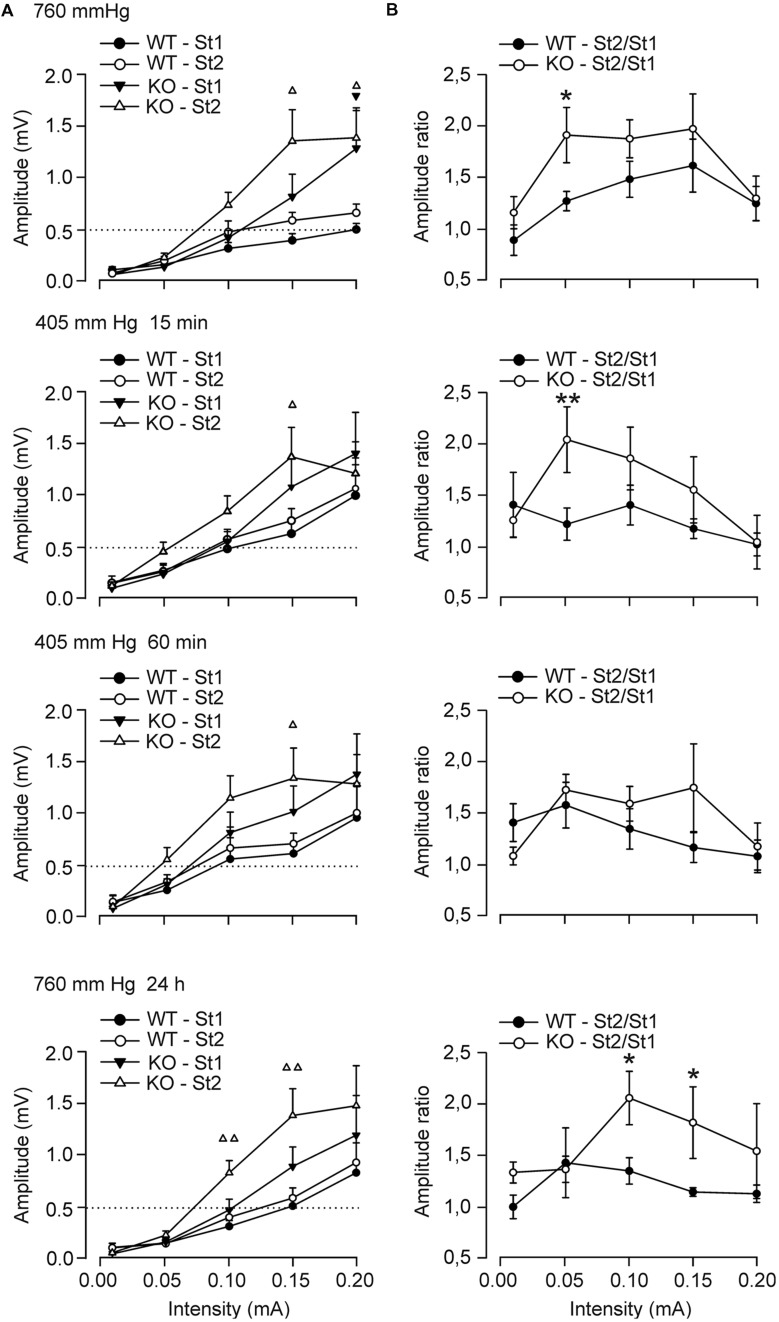
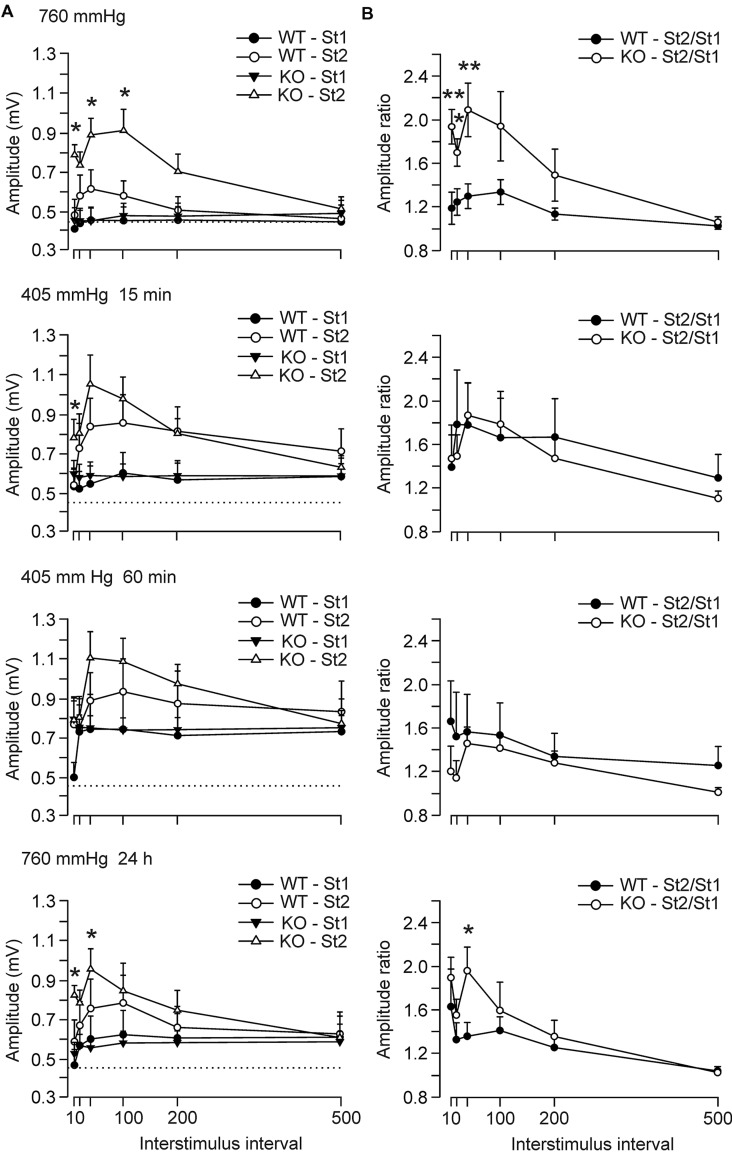
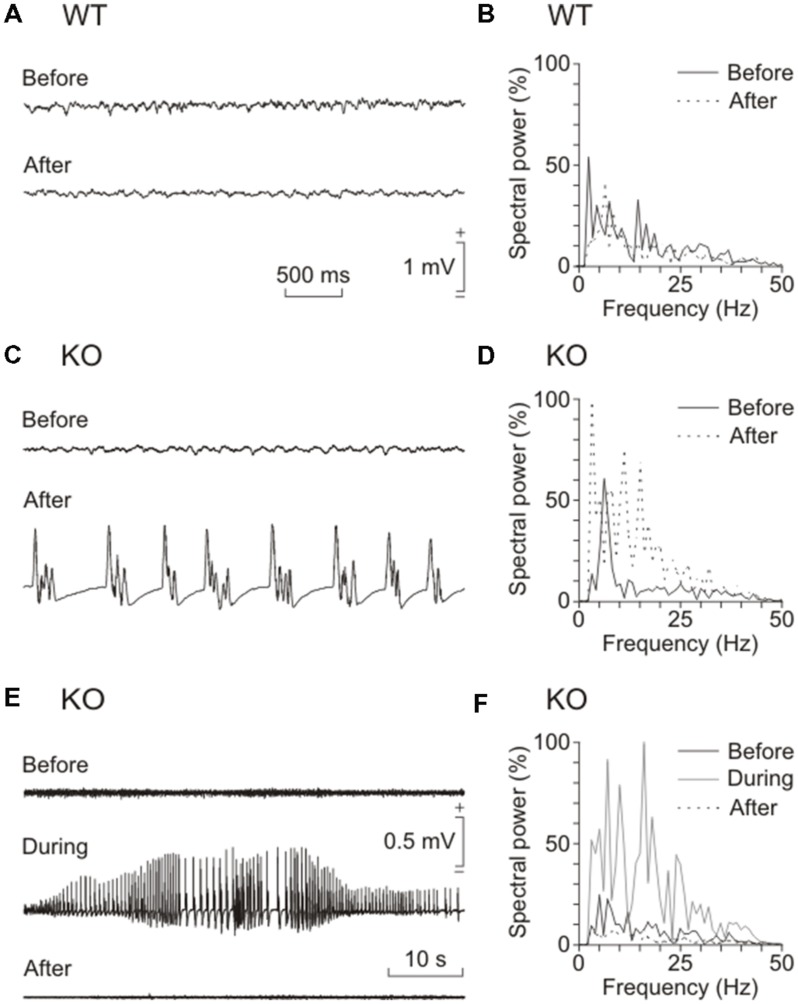
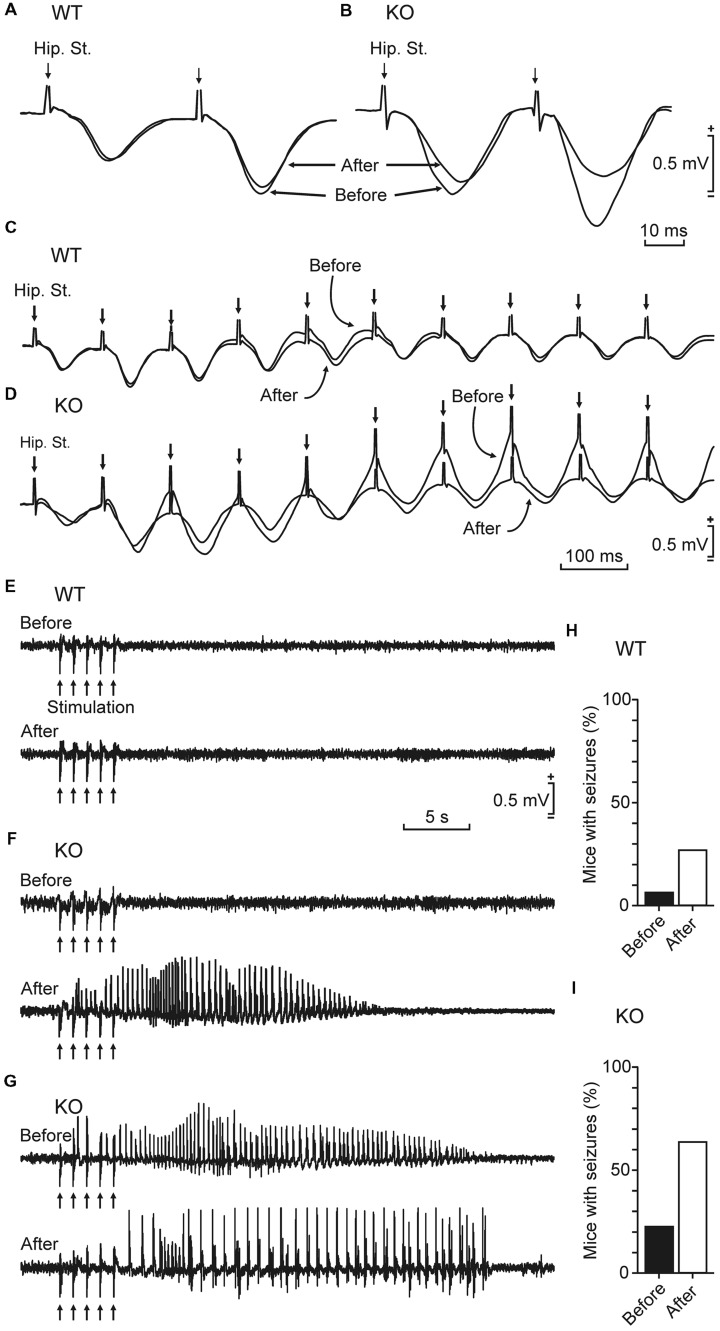
Although glycogen is the only carbohydrate reserve of the brain, its overall contribution to brain functions remains unclear. It has been proposed that glycogen participates in the preservation of such functions during hypoxia. Several reports also describe a relationship between brain glycogen and susceptibility to epilepsy. To address these issues, we used our brain-specific Glycogen Synthase knockout (GYS1(Nestin-KO)) mouse to study the functional consequences of glycogen depletion in the brain under hypoxic conditions and susceptibility to epilepsy. GYS1(Nestin-KO) mice presented significantly different power spectra of hippocampal local field potentials (LFPs) than controls under hypoxic conditions. In addition, they showed greater excitability than controls for paired-pulse facilitation evoked at the hippocampal CA3-CA1 synapse during experimentally induced hypoxia, thereby suggesting a compensatory switch to presynaptic mechanisms. Furthermore, GYS1(Nestin-KO) mice showed greater susceptibility to hippocampal seizures and myoclonus following the administration of kainate and/or a brief train stimulation of Schaffer collaterals. We conclude that brain glycogen could play a protective role both in hypoxic situations and in the prevention of brain seizures.
Keywords: brain glycogen; epilepsy; hypobaric hypoxia; kainate; local field potentials; mice.
Figures
Similar articles
-
Lack of Astrocytic Glycogen Alters Synaptic Plasticity but Not Seizure Susceptibility.Mol Neurobiol. 2020 Nov;57(11):4657-4666. doi: 10.1007/s12035-020-02055-5. Epub 2020 Aug 8. Mol Neurobiol. 2020. PMID: 32770452 Free PMC article.
-
Impairment in long-term memory formation and learning-dependent synaptic plasticity in mice lacking glycogen synthase in the brain.J Cereb Blood Flow Metab. 2013 Apr;33(4):550-6. doi: 10.1038/jcbfm.2012.200. Epub 2013 Jan 2. J Cereb Blood Flow Metab. 2013. PMID: 23281428 Free PMC article.
-
Lack of Neuronal Glycogen Impairs Memory Formation and Learning-Dependent Synaptic Plasticity in Mice.Front Cell Neurosci. 2019 Aug 13;13:374. doi: 10.3389/fncel.2019.00374. eCollection 2019. Front Cell Neurosci. 2019. PMID: 31456667 Free PMC article.
-
Impaired glucose metabolism and exercise capacity with muscle-specific glycogen synthase 1 (gys1) deletion in adult mice.Mol Metab. 2016 Jan 21;5(3):221-232. doi: 10.1016/j.molmet.2016.01.004. eCollection 2016 Mar. Mol Metab. 2016. PMID: 26977394 Free PMC article.
-
Glycogen in Astrocytes and Neurons: Physiological and Pathological Aspects.Adv Neurobiol. 2019;23:311-329. doi: 10.1007/978-3-030-27480-1_10. Adv Neurobiol. 2019. PMID: 31667813 Free PMC article. Review.
Cited by
-
Potential new roles for glycogen in epilepsy.Epilepsia. 2023 Jan;64(1):29-53. doi: 10.1111/epi.17412. Epub 2022 Nov 1. Epilepsia. 2023. PMID: 36117414 Free PMC article. Review.
-
Gys1 Antisense Therapy Prevents Disease-Driving Aggregates and Epileptiform Discharges in a Lafora Disease Mouse Model.Neurotherapeutics. 2023 Oct;20(6):1808-1819. doi: 10.1007/s13311-023-01434-9. Epub 2023 Sep 12. Neurotherapeutics. 2023. PMID: 37700152 Free PMC article.
-
Pilocarpine-Induced Status Epilepticus Is Associated with P-Glycoprotein Induction in Cardiomyocytes, Electrocardiographic Changes, and Sudden Death.Pharmaceuticals (Basel). 2018 Feb 16;11(1):21. doi: 10.3390/ph11010021. Pharmaceuticals (Basel). 2018. PMID: 29462915 Free PMC article.
-
Neurovascular coupling is optimized to compensate for the increase in proton production from nonoxidative glycolysis and glycogenolysis during brain activation and maintain homeostasis of pH, pCO2, and pO2.J Neurochem. 2024 May;168(5):632-662. doi: 10.1111/jnc.15839. Epub 2023 Jun 21. J Neurochem. 2024. PMID: 37150946 Free PMC article.
-
Lack of Astrocytic Glycogen Alters Synaptic Plasticity but Not Seizure Susceptibility.Mol Neurobiol. 2020 Nov;57(11):4657-4666. doi: 10.1007/s12035-020-02055-5. Epub 2020 Aug 8. Mol Neurobiol. 2020. PMID: 32770452 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous