

Linkage disequilibrium in humans: models and data
- PMID: 11410837
- PMCID: PMC1226024
- DOI: 10.1086/321275
Linkage disequilibrium in humans: models and data
Abstract
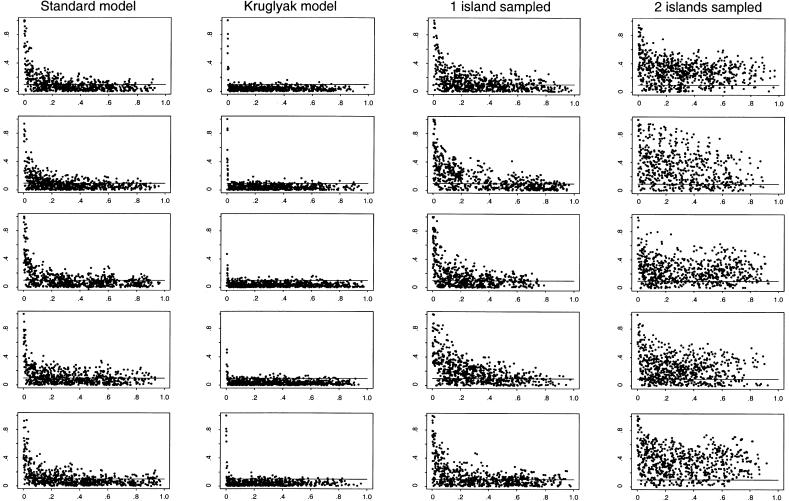
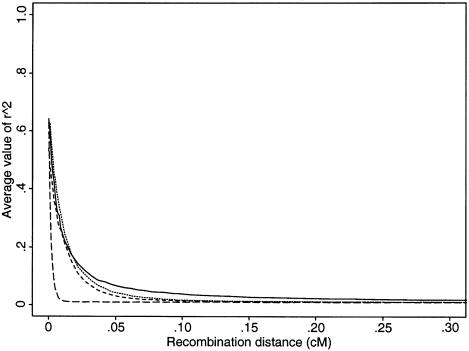
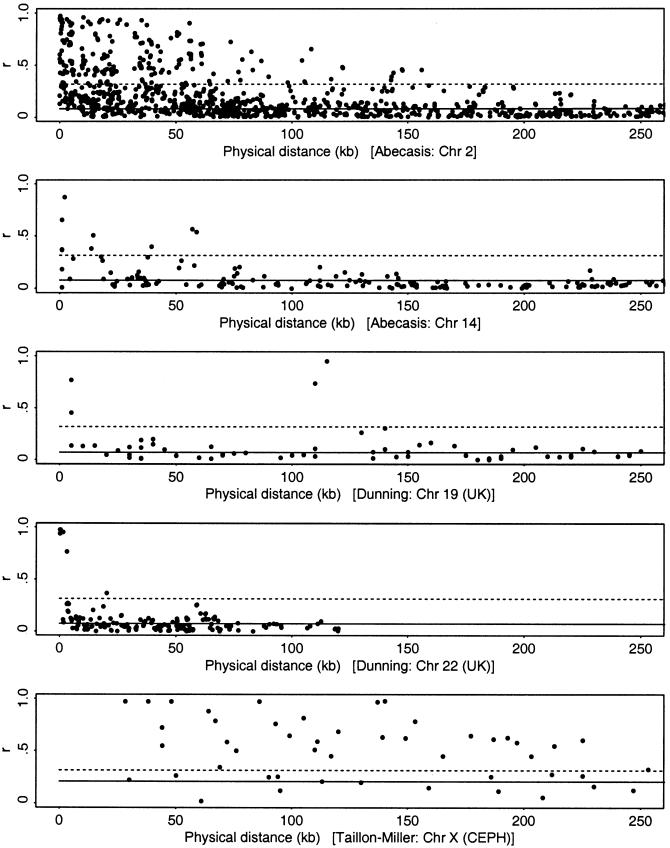
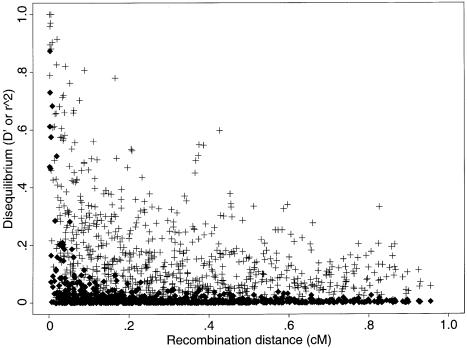
In this review, we describe recent empirical and theoretical work on the extent of linkage disequilibrium (LD) in the human genome, comparing the predictions of simple population-genetic models to available data. Several studies report significant LD over distances longer than those predicted by standard models, whereas some data from short, intergenic regions show less LD than would be expected. The apparent discrepancies between theory and data present a challenge-both to modelers and to human geneticists-to identify which important features are missing from our understanding of the biological processes that give rise to LD. Salient features may include demographic complications such as recent admixture, as well as genetic factors such as local variation in recombination rates, gene conversion, and the potential segregation of inversions. We also outline some implications that the emerging patterns of LD have for association-mapping strategies. In particular, we discuss what marker densities might be necessary for genomewide association scans.
Figures
Similar articles
-
The pattern of polymorphism on human chromosome 21.Genome Res. 2003 Jun;13(6A):1158-68. doi: 10.1101/gr.466303. Genome Res. 2003. PMID: 12799351 Free PMC article.
-
Insights into recombination from patterns of linkage disequilibrium in humans.Genetics. 2004 May;167(1):387-97. doi: 10.1534/genetics.167.1.387. Genetics. 2004. PMID: 15166163 Free PMC article.
-
Haplotype block linkage disequilibrium mapping.Front Biosci. 2003 May 1;8:a85-93. doi: 10.2741/919. Front Biosci. 2003. PMID: 12700120
-
Linkage disequilibrium for different scales and applications.Brief Bioinform. 2004 Dec;5(4):355-64. doi: 10.1093/bib/5.4.355. Brief Bioinform. 2004. PMID: 15606972 Review.
-
Mapping genes through the use of linkage disequilibrium generated by genetic drift: 'drift mapping' in small populations with no demographic expansion.Hum Hered. 1998 May-Jun;48(3):138-54. doi: 10.1159/000022794. Hum Hered. 1998. PMID: 9618061 Review.
Cited by
-
Genetic Control of Water and Nitrate Capture and Their Use Efficiency in Lettuce (Lactuca sativa L.).Front Plant Sci. 2016 Mar 30;7:343. doi: 10.3389/fpls.2016.00343. eCollection 2016. Front Plant Sci. 2016. PMID: 27064203 Free PMC article.
-
Genotype Imputation for Latinos Using the HapMap and 1000 Genomes Project Reference Panels.Front Genet. 2012 Jun 27;3:117. doi: 10.3389/fgene.2012.00117. eCollection 2012. Front Genet. 2012. PMID: 22754564 Free PMC article.
-
Potential Benefits and Challenges of Quantifying Pseudoreplication in Genomic Data with Entropy Statistics.Entropy (Basel). 2024 Sep 21;26(9):805. doi: 10.3390/e26090805. Entropy (Basel). 2024. PMID: 39330138 Free PMC article.
-
Gastroesophageal reflux disease and risk for arrhythmias: a Mendelian randomization analysis.Front Cardiovasc Med. 2024 Jul 29;11:1411784. doi: 10.3389/fcvm.2024.1411784. eCollection 2024. Front Cardiovasc Med. 2024. PMID: 39135614 Free PMC article.
-
Primary biliary cirrhosis and osteoporosis: a bidirectional two-sample Mendelian randomization study.Front Immunol. 2023 Dec 14;14:1269069. doi: 10.3389/fimmu.2023.1269069. eCollection 2023. Front Immunol. 2023. PMID: 38162659 Free PMC article.
References
Electronic-Database Information
-
- Gil McVean web site, http://www.stats.ox.ac.uk/~mcvean/ (for estimation of ρ)
-
- Hudson Lab Home Page, http://home.uchicago.edu/∼rhudson1/ (for simulation of the coalescent with recombination [see the program “mksamples”] and estimation of ρ [see the article “Two locus sampling distributions and their application”)
-
- Human Recombination Rates web site, http://eebweb.arizona.edu/nachman/publications/data/microsats.html (for estimation of c)
-
- Nordborg web site, http://www-hto.usc.edu/people/nordborg/papers.html (for the article “Linkage disequilibrium, haplotype evolution and the coalescent”)
-
- Paul Fearnhead web site, http://www.stats.ox.ac.uk/∼fhead/index.html (for estimation of ρ [see the article “Estimating recombination rates from population genetic data”])
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
