

Autophosphorylation-dependent remodeling of the DNA-dependent protein kinase catalytic subunit regulates ligation of DNA ends
- PMID: 15314205
- PMCID: PMC514382
- DOI: 10.1093/nar/gkh761
Autophosphorylation-dependent remodeling of the DNA-dependent protein kinase catalytic subunit regulates ligation of DNA ends
Abstract
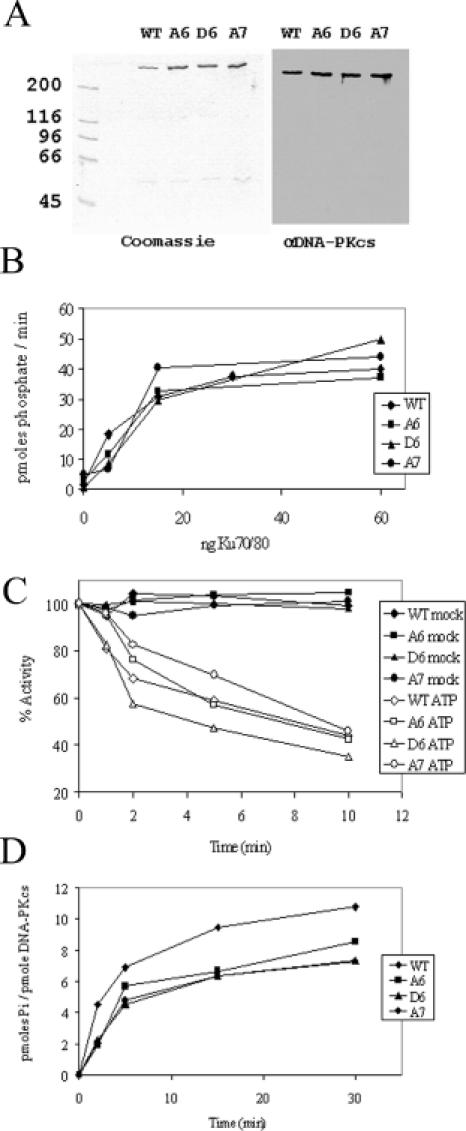
Non-homologous end joining (NHEJ) is one of the primary pathways for the repair of ionizing radiation (IR)-induced DNA double-strand breaks (DSBs) in mammalian cells. Proteins required for NHEJ include the catalytic subunit of the DNA-dependent protein kinase (DNA-PKcs), Ku, XRCC4 and DNA ligase IV. Current models predict that DNA-PKcs, Ku, XRCC4 and DNA ligase IV assemble at DSBs and that the protein kinase activity of DNA-PKcs is essential for NHEJ-mediated repair of DSBs in vivo. We previously identified a cluster of autophosphorylation sites between amino acids 2609 and 2647 of DNA-PKcs. Cells expressing DNA-PKcs in which these autophosphorylation sites have been mutated to alanine are highly radiosensitive and defective in their ability to repair DSBs in the context of extrachromosomal assays. Here, we show that cells expressing DNA-PKcs with mutated autophosphorylation sites are also defective in the repair of IR-induced DSBs in the context of chromatin. Purified DNA-PKcs proteins containing serine/threonine to alanine or aspartate mutations at this cluster of autophosphorylation sites were indistinguishable from wild-type (wt) protein with respect to protein kinase activity. However, mutant DNA-PKcs proteins were defective relative to wt DNA-PKcs with respect to their ability to support T4 DNA ligase-mediated intermolecular ligation of DNA ends. We propose that autophosphorylation of DNA-PKcs at this cluster of sites is important for remodeling of DNA-PK complexes at DNA ends prior to DNA end joining.
Figures




Similar articles
-
Analysis of DNA-dependent protein kinase-mediated DNA end joining by two-photon fluorescence cross-correlation spectroscopy.Biochemistry. 2006 Apr 4;45(13):4164-72. doi: 10.1021/bi0524060. Biochemistry. 2006. PMID: 16566590
-
Mechanisms of DNA double strand break repair and chromosome aberration formation.Cytogenet Genome Res. 2004;104(1-4):14-20. doi: 10.1159/000077461. Cytogenet Genome Res. 2004. PMID: 15162010 Review.
-
Non-homologous end joining requires that the DNA-PK complex undergo an autophosphorylation-dependent rearrangement at DNA ends.J Biol Chem. 2004 Sep 17;279(38):39408-13. doi: 10.1074/jbc.M406432200. Epub 2004 Jul 15. J Biol Chem. 2004. PMID: 15258142
-
Biochemical evidence for Ku-independent backup pathways of NHEJ.Nucleic Acids Res. 2003 Sep 15;31(18):5377-88. doi: 10.1093/nar/gkg728. Nucleic Acids Res. 2003. PMID: 12954774 Free PMC article.
-
The life and death of DNA-PK.Oncogene. 2005 Feb 3;24(6):949-61. doi: 10.1038/sj.onc.1208332. Oncogene. 2005. PMID: 15592499 Review.
Cited by
-
trans Autophosphorylation at DNA-dependent protein kinase's two major autophosphorylation site clusters facilitates end processing but not end joining.Mol Cell Biol. 2007 May;27(10):3881-90. doi: 10.1128/MCB.02366-06. Epub 2007 Mar 12. Mol Cell Biol. 2007. PMID: 17353268 Free PMC article.
-
KU-0060648 inhibits hepatocellular carcinoma cells through DNA-PKcs-dependent and DNA-PKcs-independent mechanisms.Oncotarget. 2016 Mar 29;7(13):17047-59. doi: 10.18632/oncotarget.7742. Oncotarget. 2016. PMID: 26933997 Free PMC article.
-
Structure and function of the apical PIKKs in double-strand break repair.Curr Opin Struct Biol. 2023 Oct;82:102651. doi: 10.1016/j.sbi.2023.102651. Epub 2023 Jul 10. Curr Opin Struct Biol. 2023. PMID: 37437397 Free PMC article. Review.
-
Terminal DNA structure and ATP influence binding parameters of the DNA-dependent protein kinase at an early step prior to DNA synapsis.Nucleic Acids Res. 2006 Feb 18;34(4):1112-20. doi: 10.1093/nar/gkj504. Print 2006. Nucleic Acids Res. 2006. PMID: 16488883 Free PMC article.
-
Extent to which hairpin opening by the Artemis:DNA-PKcs complex can contribute to junctional diversity in V(D)J recombination.Nucleic Acids Res. 2007;35(20):6917-23. doi: 10.1093/nar/gkm823. Epub 2007 Oct 11. Nucleic Acids Res. 2007. PMID: 17932067 Free PMC article.
References
-
- West S.C. (2003) Molecular views of recombination proteins and their control. Nature Rev. Mol. Cell Biol., 4, 435–445. - PubMed
-
- Lees-Miller S.P. and Meek,K. (2003) Repair of DNA double strand breaks by non-homologous end joining. Biochimie, 85, 1161–1173. - PubMed
-
- Valerie K. and Povirk,L.F. (2003) Regulation and mechanisms of mammalian double-strand break repair. Oncogene, 22, 5792–5812. - PubMed
-
- Lieber M.R., Ma,Y., Pannicke,U. and Schwarz,K. (2003) Mechanism and regulation of human non-homologous DNA end-joining. Nature Rev. Mol. Cell Biol., 4, 712–720. - PubMed
-
- O'Neill T., Dwyer,A.J., Ziv,Y., Chan,D.W., Lees-Miller,S.P., Abraham,R.H., Lai,J.H., Hill,D., Shiloh,Y., Cantley,L.C. and Rathbun,G.A. (2000) Utilization of oriented peptide libraries to identify substrate motifs selected by ATM. J. Biol. Chem., 275, 22719–22727. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
