

Morphine induces ubiquitin-proteasome activity and glutamate transporter degradation
- PMID: 18539596
- PMCID: PMC2490772
- DOI: 10.1074/jbc.M800809200
Morphine induces ubiquitin-proteasome activity and glutamate transporter degradation
Abstract
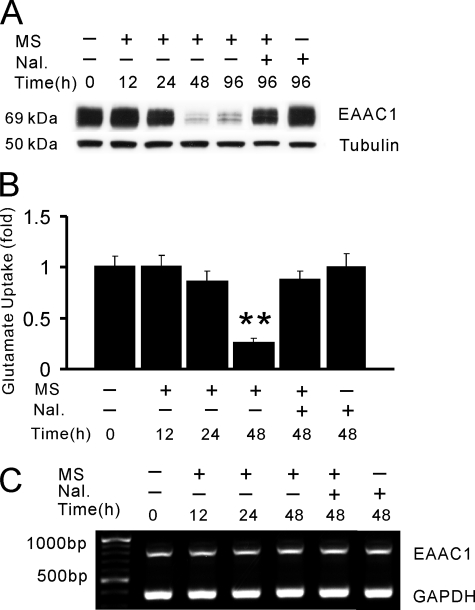
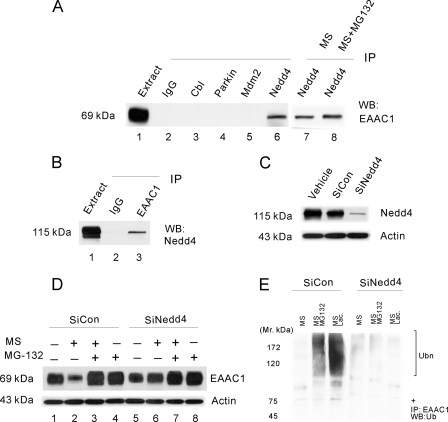
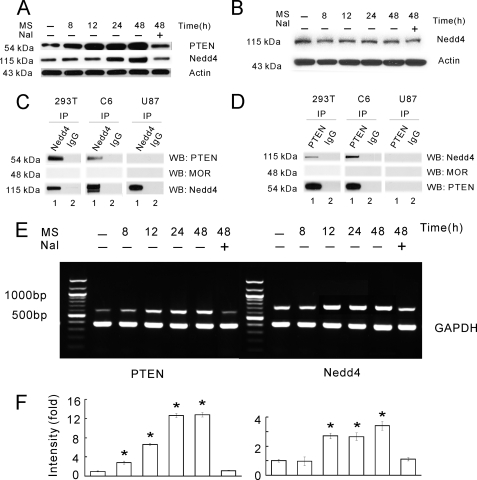
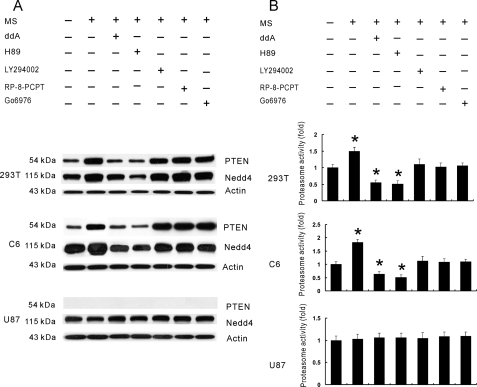
Glutamate transporters play a crucial role in physiological glutamate homeostasis, neurotoxicity, and glutamatergic regulation of opioid tolerance. However, how the glutamate transporter turnover is regulated remains poorly understood. Here we show that chronic morphine exposure induced posttranscriptional down-regulation of the glutamate transporter EAAC1 in C6 glioma cells with a concurrent decrease in glutamate uptake and increase in proteasome activity, which were blocked by the selective proteasome inhibitor MG-132 or lactacystin but not the lysosomal inhibitor chloroquin. At the cellular level, chronic morphine induced the PTEN (phosphatase and tensin homolog deleted on chromosome Ten)-mediated up-regulation of the ubiquitin E3 ligase Nedd4 via cAMP/protein kinase A signaling, leading to EAAC1 ubiquitination and proteasomal degradation. Either Nedd4 or PTEN knockdown with small interfering RNA prevented the morphine-induced EAAC1 degradation and decreased glutamate uptake. These data indicate that cAMP/protein kinase A signaling serves as an intracellular regulator upstream to the activation of the PTEN/Nedd4-mediated ubiquitin-proteasome system activity that is critical for glutamate transporter turnover. Under an in vivo condition, chronic morphine exposure also induced posttranscriptional down-regulation of the glutamate transporter EAAC1, which was prevented by MG-132, and transcriptional up-regulation of PTEN and Nedd4 within the spinal cord dorsal horn. Thus, inhibition of the ubiquitin-proteasome-mediated glutamate transporter degradation may be an important mechanism for preventing glutamate overexcitation and may offer a new strategy for treating certain neurological disorders and improving opioid therapy in chronic pain management.
Figures



Similar articles
-
Inhibition of the ubiquitin-proteasome activity prevents glutamate transporter degradation and morphine tolerance.Pain. 2008 Dec;140(3):472-478. doi: 10.1016/j.pain.2008.09.028. Epub 2008 Nov 4. Pain. 2008. PMID: 18986766 Free PMC article.
-
cAMP and protein kinase A contribute to the downregulation of spinal glutamate transporters after chronic morphine.Neurosci Lett. 2005 Mar 7;376(1):9-13. doi: 10.1016/j.neulet.2004.11.016. Epub 2004 Dec 2. Neurosci Lett. 2005. PMID: 15694265
-
Chronic morphine induces downregulation of spinal glutamate transporters: implications in morphine tolerance and abnormal pain sensitivity.J Neurosci. 2002 Sep 15;22(18):8312-23. doi: 10.1523/JNEUROSCI.22-18-08312.2002. J Neurosci. 2002. PMID: 12223586 Free PMC article.
-
Role of the ubiquitin-proteasome system in brain ischemia: friend or foe?Prog Neurobiol. 2014 Jan;112:50-69. doi: 10.1016/j.pneurobio.2013.10.003. Epub 2013 Oct 22. Prog Neurobiol. 2014. PMID: 24157661 Review.
-
Involvement of glial glutamate transporters in morphine dependence.Ann N Y Acad Sci. 2004 Oct;1025:383-8. doi: 10.1196/annals.1307.047. Ann N Y Acad Sci. 2004. PMID: 15542740 Review.
Cited by
-
Lipophilic analogues of D-cysteine prevent and reverse physical dependence to fentanyl in male rats.Front Pharmacol. 2024 Apr 5;14:1336440. doi: 10.3389/fphar.2023.1336440. eCollection 2023. Front Pharmacol. 2024. PMID: 38645835 Free PMC article.
-
Morphine induces redox-based changes in global DNA methylation and retrotransposon transcription by inhibition of excitatory amino acid transporter type 3-mediated cysteine uptake.Mol Pharmacol. 2014 May;85(5):747-57. doi: 10.1124/mol.114.091728. Epub 2014 Feb 25. Mol Pharmacol. 2014. PMID: 24569088 Free PMC article.
-
Tert-butylhydroquinone lowers blood pressure in AngII-induced hypertension in mice via proteasome-PTEN-Akt-eNOS pathway.Sci Rep. 2016 Jul 20;6:29589. doi: 10.1038/srep29589. Sci Rep. 2016. Retraction in: Sci Rep. 2019 Nov 19;9(1):17122. doi: 10.1038/s41598-019-53022-7. PMID: 27435826 Free PMC article. Retracted.
-
SLC25A22 Promotes Proliferation and Metastasis of Osteosarcoma Cells via the PTEN Signaling Pathway.Technol Cancer Res Treat. 2018 Jan 1;17:1533033818811143. doi: 10.1177/1533033818811143. Technol Cancer Res Treat. 2018. PMID: 30482097 Free PMC article.
-
Non-canonical Wnt signaling induces ubiquitination and degradation of Syndecan4.J Biol Chem. 2010 Sep 17;285(38):29546-55. doi: 10.1074/jbc.M110.155812. Epub 2010 Jul 16. J Biol Chem. 2010. PMID: 20639201 Free PMC article.
References
-
- Lievens, J. C., Bernal, F., Forni, C., Mahy, N., and Kerkerian-Le Goff, L. (2000) Glia 29 222-232 - PubMed
-
- Trotti, D., Aoki, M., Pasinelli, P., Berger, U. V., Danbolt, N. C., Brown, R. H., Jr., and Hediger, M. A. (2001) J. Biol. Chem. 276 576-582 - PubMed
-
- Vorwerk, C. K., Naskar, R., Schuettauf, F., Quinto, K., Zurakowski, D., Gochenauer, G., Robinson, M. B., Mackler, S. A., and Dreyer, E. B. (2000) Investig. Ophthalmol. Vis. Sci. 41 3615-3621 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
