

Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis
- PMID: 24412617
- PMCID: PMC4305274
- DOI: 10.1016/j.immuni.2013.12.007
Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis
Abstract
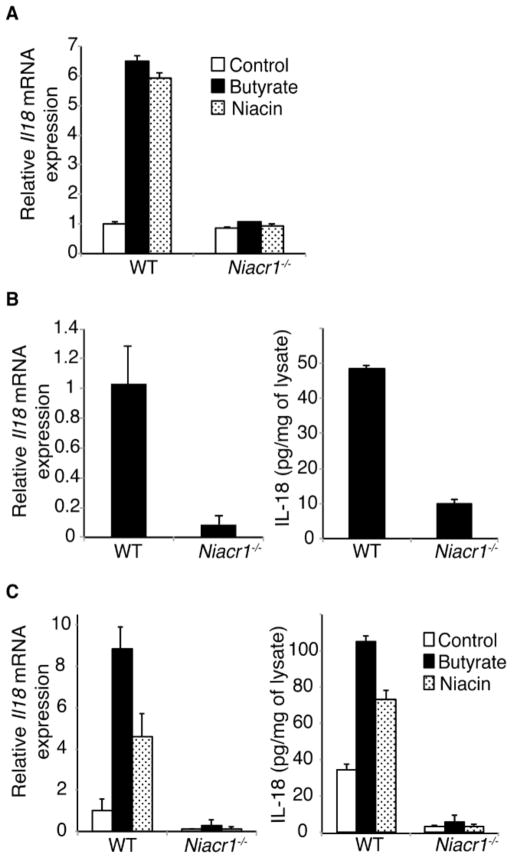
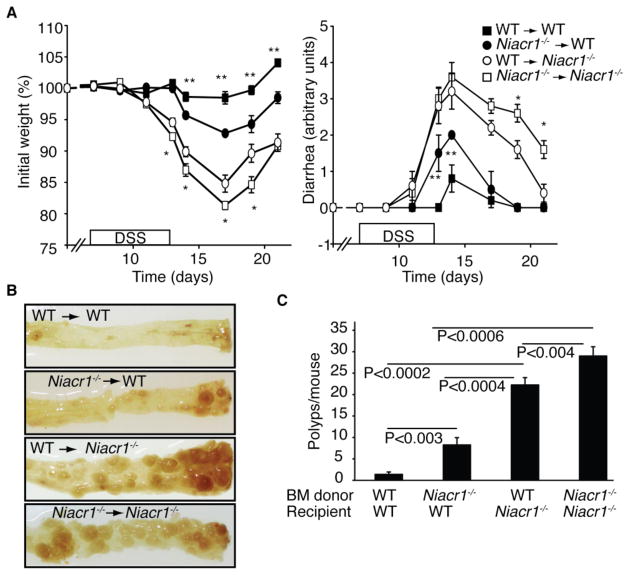
Commensal gut microflora and dietary fiber protect against colonic inflammation and colon cancer through unknown targets. Butyrate, a bacterial product from fermentation of dietary fiber in the colon, has been implicated in this process. GPR109A (encoded by Niacr1) is a receptor for butyrate in the colon. GPR109A is also a receptor for niacin, which is also produced by gut microbiota and suppresses intestinal inflammation. Here we showed that Gpr109a signaling promoted anti-inflammatory properties in colonic macrophages and dendritic cells and enabled them to induce differentiation of Treg cells and IL-10-producing T cells. Moreover, Gpr109a was essential for butyrate-mediated induction of IL-18 in colonic epithelium. Consequently, Niacr1(-/-) mice were susceptible to development of colonic inflammation and colon cancer. Niacin, a pharmacological Gpr109a agonist, suppressed colitis and colon cancer in a Gpr109a-dependent manner. Thus, Gpr10a has an essential role in mediating the beneficial effects of gut microbiota and dietary fiber in colon.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures





Comment in
-
GPR109a: the missing link between microbiome and good health?Immunity. 2014 Jan 16;40(1):8-10. doi: 10.1016/j.immuni.2013.12.009. Immunity. 2014. PMID: 24439263 Free PMC article.
Similar articles
-
GPR109a: the missing link between microbiome and good health?Immunity. 2014 Jan 16;40(1):8-10. doi: 10.1016/j.immuni.2013.12.009. Immunity. 2014. PMID: 24439263 Free PMC article.
-
GPR109A is a G-protein-coupled receptor for the bacterial fermentation product butyrate and functions as a tumor suppressor in colon.Cancer Res. 2009 Apr 1;69(7):2826-32. doi: 10.1158/0008-5472.CAN-08-4466. Epub 2009 Mar 10. Cancer Res. 2009. PMID: 19276343 Free PMC article.
-
Gpr109a Limits Microbiota-Induced IL-23 Production To Constrain ILC3-Mediated Colonic Inflammation.J Immunol. 2018 Apr 15;200(8):2905-2914. doi: 10.4049/jimmunol.1701625. Epub 2018 Mar 7. J Immunol. 2018. PMID: 29514953 Free PMC article.
-
Benefits of short-chain fatty acids and their receptors in inflammation and carcinogenesis.Pharmacol Ther. 2016 Aug;164:144-51. doi: 10.1016/j.pharmthera.2016.04.007. Epub 2016 Apr 23. Pharmacol Ther. 2016. PMID: 27113407 Free PMC article. Review.
-
A novel treatment target for Parkinson's disease.J Neurol Sci. 2014 Dec 15;347(1-2):34-8. doi: 10.1016/j.jns.2014.10.024. Epub 2014 Oct 23. J Neurol Sci. 2014. PMID: 25455298 Review.
Cited by
-
Microbiota and COVID-19: Long-term and complex influencing factors.Front Microbiol. 2022 Aug 12;13:963488. doi: 10.3389/fmicb.2022.963488. eCollection 2022. Front Microbiol. 2022. PMID: 36033885 Free PMC article. Review.
-
Immunity and Tolerance Induced by Intestinal Mucosal Dendritic Cells.Mediators Inflamm. 2016;2016:3104727. doi: 10.1155/2016/3104727. Epub 2016 Feb 29. Mediators Inflamm. 2016. PMID: 27034589 Free PMC article. Review.
-
The gut microbiota associated with high-Gleason prostate cancer.Cancer Sci. 2021 Aug;112(8):3125-3135. doi: 10.1111/cas.14998. Epub 2021 Jul 5. Cancer Sci. 2021. PMID: 34051009 Free PMC article.
-
Microbiota metabolite regulation of host immune homeostasis: a mechanistic missing link.Curr Allergy Asthma Rep. 2015 May;15(5):24. doi: 10.1007/s11882-015-0524-2. Curr Allergy Asthma Rep. 2015. PMID: 26139332 Review.
-
The Microbiome-Gut-Brain Axis and Resilience to Developing Anxiety or Depression under Stress.Microorganisms. 2021 Mar 31;9(4):723. doi: 10.3390/microorganisms9040723. Microorganisms. 2021. PMID: 33807290 Free PMC article. Review.
References
-
- Backhed F, Ley RE, Sonnenburg JL, Peterson DA, Gordon JI. Host-bacterial mutualism in the human intestine. Science. 2005;307:1915–1920. - PubMed
-
- Blad CC, Tang C, Offermanns S. G protein-coupled receptors for energy metabolites as new therapeutic targets. Nat Rev Drug Discov. 2012;11:603–619. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
