

Heat transduction in rat sensory neurons by calcium-dependent activation of a cation channel
- PMID: 9192682
- PMCID: PMC21275
- DOI: 10.1073/pnas.94.13.7006
Heat transduction in rat sensory neurons by calcium-dependent activation of a cation channel
Abstract
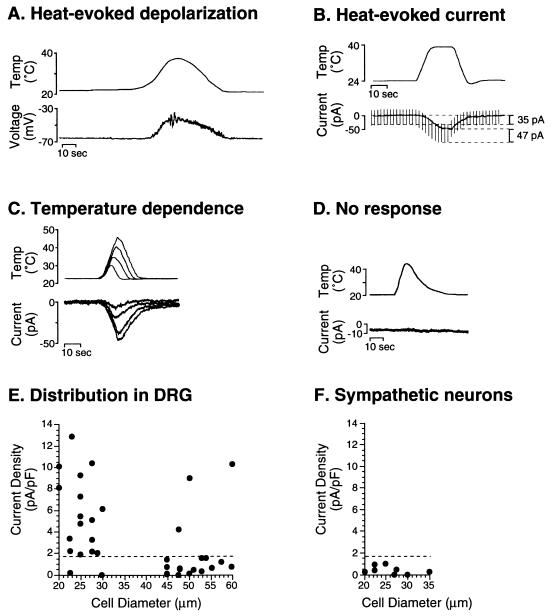
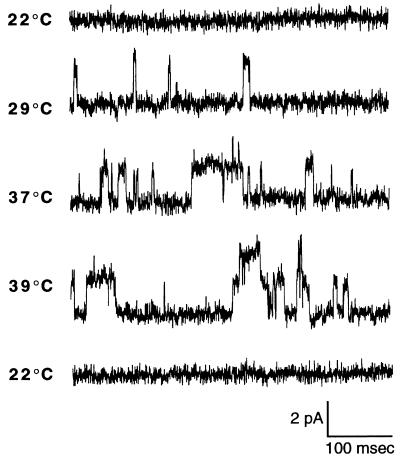
The mechanism of heat transduction in vertebrate sensory neurons was investigated in vitro by using cultured dorsal root ganglion neurons from adult rat. In response to a physiologically relevant range of stimulus temperatures (23-45 degrees C), a subpopulation of small dorsal root ganglion neurons are depolarized by a cation current (heat-activated current, Iheat) that is antagonized by extracellular cesium. Heat-induced single-channel currents in cell-attached patches are evoked at a similar range of temperatures. Iheat is a calcium-dependent current activated indirectly by heat-evoked release of calcium from intracellular stores. This suggests that the channel itself is not the transducer of thermal energy. Similar to nociceptive heat sensation in vivo, Iheat is enhanced by the hyperalgesic agent prostaglandin E2 and only partially adapts during prolonged heat stimuli. To our knowledge, these data provide the first demonstration that ion channels can mediate heat transduction in mammalian sensory neurons and provide evidence that heat causes the channels to open via an increase in the intracellular second messenger calcium.
Figures



Similar articles
-
Inhibition of rapid heat responses in nociceptive primary sensory neurons of rats by vanilloid receptor antagonists.J Neurophysiol. 1999 Dec;82(6):2853-60. doi: 10.1152/jn.1999.82.6.2853. J Neurophysiol. 1999. PMID: 10601424
-
Temperature coefficient of membrane currents induced by noxious heat in sensory neurones in the rat.J Physiol. 1999 May 15;517 ( Pt 1)(Pt 1):181-92. doi: 10.1111/j.1469-7793.1999.0181z.x. J Physiol. 1999. PMID: 10226158 Free PMC article.
-
Phosphorylation of extracellular signal-related protein kinase is required for rapid facilitation of heat-induced currents in rat dorsal root ganglion neurons.Neuroscience. 2006 Nov 17;143(1):253-63. doi: 10.1016/j.neuroscience.2006.07.047. Epub 2006 Sep 14. Neuroscience. 2006. PMID: 16973292
-
Sensation is painless.Trends Neurosci. 2003 Dec;26(12):643-5. doi: 10.1016/j.tins.2003.09.013. Trends Neurosci. 2003. PMID: 14624845 Review.
-
Thermoreceptors: recent heat in thermosensation.Curr Biol. 1997 Nov 1;7(11):R679-81. doi: 10.1016/s0960-9822(06)00354-x. Curr Biol. 1997. PMID: 9382796 Review.
Cited by
-
Ion channels gated by heat.Proc Natl Acad Sci U S A. 1999 Jul 6;96(14):7658-63. doi: 10.1073/pnas.96.14.7658. Proc Natl Acad Sci U S A. 1999. PMID: 10393876 Free PMC article. Review.
-
Nociceptors for the 21st century.Curr Rev Pain. 2000;4(6):451-8. doi: 10.1007/s11916-000-0069-4. Curr Rev Pain. 2000. PMID: 11060591 Review.
-
Bradykinin-12-lipoxygenase-VR1 signaling pathway for inflammatory hyperalgesia.Proc Natl Acad Sci U S A. 2002 Jul 23;99(15):10150-5. doi: 10.1073/pnas.152002699. Epub 2002 Jul 3. Proc Natl Acad Sci U S A. 2002. PMID: 12097645 Free PMC article.
-
Indirect actions of bradykinin on neonatal rat dorsal root ganglion neurones: a role for non-neuronal cells as nociceptors.J Physiol. 2001 Oct 1;536(Pt 1):111-21. doi: 10.1111/j.1469-7793.2001.00111.x. J Physiol. 2001. PMID: 11579161 Free PMC article.
-
Single-channel properties of native and cloned rat vanilloid receptors.J Physiol. 2002 Nov 15;545(1):107-17. doi: 10.1113/jphysiol.2002.016352. J Physiol. 2002. PMID: 12433953 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
